ISSN 1981-8203
Pombal, Paraíba, Brasil
doi: 10.18378/rvads.v21i1.11340

|
Revista Verde ISSN 1981-8203 Pombal, Paraíba, Brasil |
v. 21, n.1, jan.-mar., p.24-32, 2026 doi: 10.18378/rvads.v21i1.11340 |
|
Bruna Macedo Simões Sergio¹; Eyko Nathan Silva Sanches²; Yugo Lima-Melo³; Leandro Galon⁴; Alfredo Castamann⁵; Marieli Nandra Perkuhn⁶; Denise Cargnelutti⁷
Recebido: 17-11-2025
Aprovado: 02-02-2026
O presente estudo objetivou avaliar as respostas bioquímicas de plantas de milho crioulo submetidas ao déficit hídrico, adubadas com fosfato natural reativo (fnr) e inoculadas com microrganismos promotores de crescimento de plantas (mpcp). O experimento foi conduzido em vasos, em delineamento inteiramente casualizado, em esquema fatorial 2 × 7, com cinco repetições por tratamento. O primeiro fator correspondeu às condições hídricas: 100% da capacidade do vaso (cp), caracterizando ausência de déficit hídrico, e 50% da cp, caracterizando déficit hídrico. O segundo fator foi composto por sete tratamentos: controle (sem inoculação e sem fnr), fnr isolado, azospirillum brasilense (az), bacillus subtilis + bacillus megaterium (bsbm), microrganismos eficientes (me), fnr + az, e fnr + bsbm. Os microrganismos foram aplicados nas sementes antes da semeadura e diretamente no solo e nas plantas, com periodicidade semanal até 35 dias após a semeadura (das). Aos 35 das, no estádio v6, foram determinados os teores de proteínas, a atividade da enzima guaiacol peroxidase (gpod), prolina e malondialdeído (mda) nas folhas e raízes. Os teores médios de proteínas nas raízes das plantas submetidas ao déficit hídrico foram 66% superiores aos teores das plantas não submetidas ao déficit hídrico. O déficit hídrico incrementou a atividade da gpod, mas reduziu os teores de prolina e mda nas raízes. O maior acúmulo de prolina foi observado em plantas que receberam fnr+bsbm, porém, sob déficit hídrico, o tratamento bsbm induziu maior acúmulo de prolina nas folhas (3,7 vezes). Os níveis de mda (produto da peroxidação lipídica) foram maiores nas folhas no controle, az, bsbm ou fnr+bsbm de plantas sem déficit hídrico. Os bioinsumos foram capazes de amenizar as alterações bioquímicas provocadas pelo déficit hídrico em plantas de milho crioulo.
Palavras-chave: Agroecologia. Biodiversidade. Espécies reativas de oxigênio. Microrganismos eficientes. Zea mays.
The present study aimed to evaluate the biochemical responses of creole maize plants subjected to water deficit (wd), fertilized with reactive natural phosphate (rnp), and inoculated with plant growth-promoting microorganisms (pgpm). The experiment was conducted in pots in a completely randomized design, arranged in a 2 × 7 factorial scheme, with five replicates per treatment. The first factor corresponded to water conditions: 100% of pot capacity (pc), characterizing absence of water deficit, and 50% of pc, characterizing water deficit (wd). The second factor consisted of seven treatments: control (without inoculation and without RNP), RNP alone, Azospirillum brasilense (Az), Bacillus subtilis + Bacillus megaterium (BsBm), efficient microorganisms (em), rnp + Az, and rnp + BsBm. The microorganisms were applied to the seeds before sowing and directly to the soil and plants on a weekly basis until 35 days after sowing (das). At 35 DAS, at the V6 stage, protein contents, guaiacol peroxidase (gpod) enzyme activity, proline, and malondialdehyde (mda) were determined in leaves and roots. The average protein contents in the roots of plants subjected to wd were 66% higher than those of plants not subjected to wd. wd increased gpod activity but reduced proline and mda contents in the roots. The highest proline accumulation was observed in plants that received rnp+BsBm; however, under wd, the BsBm treatment induced greater proline accumulation in the leaves (3.7-fold). mda levels (a product of lipid peroxidation) were higher in the leaves of control, Az, BsBm, or rnp+BsBm plants without wd. The bioinputs were able to mitigate the biochemical alterations caused by wd in creole maize plants.
Keywords: Agroecology. Biodiversity. Reactive oxygen species. Efficient microorganisms. Zea mays.
As condições ambientais afetam o desenvolvimento das plantas, pois estas estão sujeitas a diversos estresses abióticos e bióticos, que se intensificam ao se somarem. Dentre estes, o déficit hídrico é um limitador do desenvolvimento de vegetais, pois a água é fundamental para funções metabólicas e manutenção da turgidez celular adequada. Assim, o aumento da temperatura somado à baixa regularidade de precipitação pode provocar o déficit hídrico nos vegetais, afetando o desenvolvimento de frutos e sementes (KUMARI et al., 2024).
Estudos têm mostrado uma tendência de incremento da temperatura global com consequente aumento de regiões áridas, maior periodicidade de eventos de déficit hídrico, altas temperaturas, toxicidade de nutrientes e salinidade (SARDANS et al., 2024). Essas alterações implicam direta ou indiretamente em áreas de plantio e em questões ambientais e socioeconômicas (VIEIRA et al., 2023). Fenômenos atmosféricos como El Niño e La Ninã tendem a se intensificar e provocar períodos de estiagem e aumentar a intensidade de chuvas nas regiões afetadas (GENG et al., 2023).
Dentre as culturas de relevância no mundo e no Brasil está o milho (Zea mays L.) tanto no aspecto de segurança alimentar quanto da nutrição animal e humana (CONTINI et al., 2019). Trata-se de uma das principais fontes de renda dos agricultores familiares, de acordo com o boletim de safra brasileira do ano de 2023 (CONAB, 2023).
O milho demonstra elevada sensibilidade ao estresse hídrico, o que compromete significativamente seu crescimento ao desencadear desequilíbrios nutricionais e fisiológicos (ABDELAAL et al., 2021). A falta de água reduz a disponibilidade de minerais no solo e limita o seu transporte e absorção pelos tecidos, dificultando a assimilação de elementos como o fósforo (P), essencial para processos como fotossíntese e divisão celular (ABDELAAL et al., 2021). Embora os teores totais de P nos solos agrícolas variem de 200 a 3.000 mg Kg-1 de solo, a concentração de P inorgânico na solução do solo varia de 0,1 a 10 µmol L-1 (NOVAIS et al., 2007). Menos de 0,1% do P total presente nos solos está disponível na solução para absorção pelas raízes das plantas. Consequentemente, as plantas absorvem este nutriente de soluções com baixas concentrações (SOUSA et al., 2010). Nos cultivos convencionais, geralmente aplicam-se adubos fosfatados em quantidades superiores às necessidades das plantas, sendo de fontes que possuem alto custo de importação.
Desta forma, o fosfato natural (rocha fosfática) constitui uma fonte alternativa de fósforo com eficiência dependente da reatividade do material, do tipo de solo (especialmente solos ácidos) e do manejo, podendo, em condições favoráveis, apresentar desempenho comparável a fontes solúveis e atender sistemas orgânicos e agroecológicos (HELLUMS et al., 2026). Além disso, comparações experimentais entre superfosfato e rocha fosfática indicam que a eficiência e a disponibilidade de P podem variar conforme o sistema de manejo (orgânico vs. convencional) e as características do solo, sustentando o uso do fosfato natural como fonte alternativa em contextos específicos (LENGNICK; KING, 2009).
Entre as estratégias utilizadas para o cultivo de plantas sob condições de estresse abiótico destaca-se a aplicação de microrganismos promotores de crescimento vegetal (Plant Growth-Promoting Microorganisms- PGPM). Estudos demonstram que as atividades de cepas microbianas adaptadas exercem um efeito positivo no desenvolvimento das plantas sob condições de déficit hídrico (ROLLI et al., 2015). A atividade enzimática e a síntese de proteínas podem ser otimizadas pela utilização desses microrganismos (ABDELAAL et al., 2021). Rizobactérias dos gêneros Bacillus e Azospirillum são comercializadas como inoculantes para culturas agrícolas, como milho, trigo, soja, feijão, cana-de-açúcar (MEENA et al., 2016; VEJAN et al., 2016) atuando como bioinsumos.
Os estresses abióticos, como a deficiência de água e de nutrientes essenciais, como o fósforo, desencadeiam alterações fisiológicas nas plantas, entre as quais se destaca o aumento na produção de espécies reativas de oxigênio (ERO). Embora produzidas naturalmente nas células vegetais, o aumento excessivo dessas espécies pode exceder a capacidade do sistema antioxidante enzimático e não enzimático, levando ao estresse oxidativo, e a danos às biomoléculas, como lipídios, DNA e proteínas (MITTLER et al., 2022).
Deste modo, o uso de microrganismos e a aplicação de fontes naturais de fósforo podem contribuir para a modulação de respostas bioquímicas associadas a tolerância ao déficit hídrico. Assim, o presente estudo teve como objetivo avaliar as respostas bioquímicas do uso de fosfato natural reativo (FNR) e de microrganismos promotores do crescimento de plantas (MPCP), em plantas de milho crioulo submetidas ao déficit hídrico.
Os experimentos foram desenvolvidos no Laboratório de Microscopia, Laboratório de Entomologia e Bioquímica e na casa de vegetação da Universidade Federal da Fronteira Sul, campus Erechim. Rio Grande do Sul, Brasil. Durante o período experimental (dezembro e janeiro de 2023), as condições climáticas médias na região de Erechim (RS) foram aproximadamente de 26–28 °C para temperatura máxima, 17 °C para temperatura mínima e 75–79% de umidade relativa do ar, com base em séries climatológicas históricas para o município. Foram utilizadas sementes de milho variedade crioula obtidas junto ao Centro de Apoio e Promoção da Agroecologia (CAPA – Erechim, RS) (Número de registro no SISGEN: A5A4971). Três sementes foram semeadas em vasos plásticos contendo 8 litros de substrato (solo + composto orgânico + areia, na proporção 1:1:1). Após 15 dias da germinação efetuou-se o raleio mantendo uma planta por vaso. O delineamento experimental utilizado foi o inteiramente casualizado arranjado em esquema fatorial 2 x 7, com 5 repetições. No fator A testou-se os déficits hídricos (50 e 100% CP) e no fator B testou-se os bioinsumos (Azospirillum, Az; Microrganismos Eficientes, ME; BiomaPhos, BsBm) isolados e combinados com o fosfato natural reativo (Tabela 1).
Tabela 1. Bioinsumos e níveis de déficit hídrico avaliados na cultura do milho.
| Tratamentos | 100% CP | 50% CP |
|---|---|---|
| Controle | Controle (100% CP) | Controle (50% CP) |
| Az | Azospirillum, 100% CP | Azospirillum, 50% CP |
| ME | ME 1:500, 100% CP | ME 1:500, 50% CP |
| BsBm | BiomaPhos, 100% CP | BiomaPhos, 50% CP |
| FNR+Az | Fosfato+Azospirillum, 100% CP | Fosfato+Azospirillum, 50% CP |
| FNR+BsBm | Fosfato+BiomaPhos, 100% CP | Fosfato+BiomaPhos, 50% CP |
| FNR+ME | Fosfato+ME 1:500, 100% CP | Fosfato+ME 1:500, 50% CP |
A capacidade de pote (CP) foi determinada pelo método gravimétrico, considerando-se a massa do conjunto vaso + solo seco e, posteriormente, a massa após saturação hídrica por capilaridade e drenagem livre. A massa de água retida a 100% da CP foi obtida pela diferença entre a massa do sistema saturado e a massa do conjunto vaso + solo seco. Para os vasos induzidos a 50% de restrição hídrica foi fornecido volume de água correspondente a 50% da água retida na condição de 100% da CP, mantendo-se constante a massa de solo. A umidade foi monitorada diariamente por pesagem dos vasos, com reposição da água evapotranspirada para manter os níveis hídricos estabelecidos inicialmente. Os potes sem as plantas foram pesados com intervalo de dois dias, para determinar a quantidade de água evaporada, sendo previamente conhecida a massa dos vasos que contém plantas e dos que não contém plantas. A reposição da água evaporada no período foi realizada com uso de regador manual, mantendo-se os vasos próximos à capacidade de pote para o tratamento controle (100% CP) e omitindo-se a irrigação em 50% para o tratamento de indução de déficit hídrico (50% CP). As plantas foram submetidas ao déficit hídrico 16 dias após a semeadura (DAS) as quais permaneceram nesta condição até o estádio V6, referente aos 35 DAS.
Os microrganismos eficientes (ME) utilizados nesta pesquisa foram obtidos e capturados de acordo com as normas do Ministério da Agricultura da Pecuária e Abastecimento, por meio das Fichas Agroecológicas (LEITE; MEIRA, 2016; SANTIAGO et al., 2022) na área experimental da UFFS, Campus Erechim (Número de registro no SISGEN: A5A4971). Os microrganismos, Azospirillum (Bioma mais®, Cepas ABV5/ABV6; UFC 2,5 X 10⁸) e Bacillus megaterium e B. subtilis (BiomaPhos®, Cepa BRM 2084; UFC 4 X 10⁹) foram adquiridos comercialmente e os MPCP, na forma de solução diluída (1:500) foram coletados, diluídos de forma artesanal manualmente (não há padronização da concentração celular (UFC/mL, apresentando uma limitação referente à ausência de quantificação de unidades formadoras de colônias (UFC)), aplicados em tratamento de sementes e borrifados 4 mL por vaso na planta e no solo, 2mL para o alvo biológico (folha) e para o alvo físico (solo). As sementes foram inoculadas separadamente com soluções dos microrganismos na diluição 1:500. As sementes permaneceram 30 minutos nesta solução e em seguida foram retiradas e deixadas para secar em temperatura ambiente por 24 horas.
As sementes foram previamente inoculadas antes da semeadura. As aplicações via solo e parte aérea foram realizadas semanalmente, sempre no período da tarde, iniciando na primeira semana após a semeadura (7 DAS) e estendendo-se até o 35º DAS. No tratamento controle foi borrifada apenas água. O fosfato natural reativo foi incorporado ao substrato na dose de 155 mg kg⁻¹, correspondendo a aproximadamente 1,24 g por vaso, considerando 8 L de substrato por unidade experimental. A dose foi estabelecida com base na análise química do solo.
Tabela 2. Composição química da amostra de substrato analisada.
| Parâmetro | Unidade | Valor |
|---|---|---|
| Umidade (65ºC) | % | 5,99 |
| Nitrogênio total (N) | % | 1,53 |
| Fósforo total (P₂O₅) | % | 1,70 |
| Potássio solúvel (K₂O) | % | 0,76 |
| Carbono Orgânico Total (COT) | % | 8,48 |
| pH (CaCl₂ 0,01M) | - | 7,85 |
| Relação C/N | - | 5,20 |
| Condutividade Elétrica (CE) | mS·cm⁻¹ | 2,15 |
Os parâmetros bioquímicos foram obtidos aos 35 DAS, no dia da última aplicação dos microrganismos, em que as plantas foram coletadas, divididas em folhas e raízes as quais foram imediatamente congeladas em nitrogênio líquido e armazenadas a -80 ºC.
A atividade da enzima guaiacol peroxidase (GPOD), foi determinada em amostras frescas de raízes e folhas de milho. Um grama de tecido fresco foi homogeneizado em 3 mL de uma solução tampão de fosfato de sódio (pH 7,8) 0,05 M, contendo 1 mM de EDTA e 2% (w/v) de polivinilpirrolidona (PVP). O homogeneizado foi centrifugado a 10.000 rpm por 20 minutos a 4 °C e o sobrenadante foi utilizado para o ensaio. A atividade das peroxidases não específicas presentes no extrato foi determinada utilizando-se o guaiacol como substrato (ZERAIK et al., 2008).
Para a mensuração do conteúdo de prolina foi utilizado o procedimento descrito por Bates et al. (1973), em que 0,4 g de folhas foram homogeneizadas em 4 mL de ácido sulfosalicílico 10% e centrifugado a 5000 x g à 4 °C durante 20 minutos. A mistura de reação foi composta por 1 mL de sobrenadante, 1 mL de solução ácida de ninhidrina (composta por ácido acético-glacial, ácido fosfórico 6 M e ninhidrina) e 1 mL de ácido acético glacial. Esta mistura foi incubada a 100 ºC por 1 hora e resfriada em banho de gelo. Foi adicionado 1 mL de tolueno seguido de agitação em vortex durante 20 segundos. A fase contendo o tolueno foi aspirada e utilizada para leitura à 520 nm.
A concentração de proteínas solúveis foi determinada utilizando-se o método de Bradford (KRUGER, [s.d.]) com uso de albumina sérica bovina como padrão. Os resultados obtidos para os níveis de proteínas foram expressos em MDA g⁻¹ de matéria fresca (MF).
A peroxidação de lipídios da membrana (TBARS) foi estimada seguindo o método das substâncias reativas ao ácido tiobarbitúrico (EL-MOSHATY et al., 1993). Amostras frescas de raízes e folhas (1 g) foram maceradas em nitrogênio líquido e homogeneizadas em 10 mL de 0,2 M de tampão citrato (pH 6,5) contendo 0,5% de Triton X-100. O homogeneizado foi centrifugado por 15 minutos a 20.000 x g. Um mL do sobrenadante foi adicionado a 1 mL de ácido tricloroacético 20% (w/v) contendo 0,5% (w/v) de ácido tiobarbitúrico. A mistura foi aquecida a 95 ºC por 40 min e então resfriada por 15 min, centrifugada a 5.000 x g por 15 min. A absorbância do sobrenadante foi lida a 532 e 600 nm (para corrigir a turbidez não específica). A peroxidação lipídica foi expressa como nmol MDA g⁻¹ de matéria fresca (MF).
Os dados foram submetidos à análise de variância (teste F) considerando as fontes de variação déficit, tratamentos e a interação déficit x tratamentos. As médias dos tratamentos foram comparadas pelo Teste de Tukey a 5% de probabilidade de erro. As análises estatísticas foram realizadas utilizando o software genes e as análises de correlações foram obtidas por meio do software GraphPad Prism 10.
A análise de variância evidenciou interação significativa entre os tratamentos e a condição hídrica para a peroxidação lipídica (TBARS) nas folhas (GL = 4; F = 0,22; p = 0,925). Para as demais variáveis avaliadas, conforme apresentado na Tabela 3, também foram observadas interações entre os fatores. Adicionalmente, para a peroxidação lipídica nas raízes, observou-se efeito significativo da condição hídrica (GL = 1; F = 11,78; p = 0,003), enquanto os efeitos dos tratamentos (GL = 4; F = 0,44; p = 0,777) e da interação entre os fatores (GL = 4; F = 0,41; p = 0,798) não foram significativos.
No tratamento ME sob 100% da capacidade de vaso, o teor de proteínas nas raízes foi de 11,34 mg g⁻¹ MF valor superior ao observado no controle na mesma condição (TC = 2,10 mg g⁻¹ MF) e inferior ao tratamento ME sob 50% (37,77mg g⁻¹ MF). Essas diferenças foram confirmadas pelo teste de Tukey (p ≤ 0,05), conforme indicado pelas letras associadas às médias (Tabela 4). A redução dos teores de proteínas solúveis pode ser atribuída ao aumento da atividade das enzimas proteolíticas, responsáveis pela degradação das proteínas de reserva das plantas (xing et al., 2023). Por outro lado, em relação às proteínas das raízes foi observado um incremento médio em plantas expostas ou não ao déficit hídrico, em todos os tratamentos testados, quando comparado com a média dos tratamentos controle (média entre 100% e 50% cp). A análise de variância indicou efeito significativo dos tratamentos (GL = 4; F = 6,31; p = 0,002), da condição hídrica (GL = 1; F = 466,02; p < 0,001) e da interação entre os fatores (GL = 4; F = 6,15; p = 0,002). De modo geral, observou-se aumento nos níveis de proteínas nas raízes em todos os tratamentos. Sob condição de déficit hídrico, foram observadas maiores concentrações de proteínas (13,59 mg g⁻¹ MF) em comparação às plantas não submetidas ao estresse (4,75 mg g⁻¹ MF), indicando que o déficit hídrico induz alterações metabólicas no milho crioulo, possivelmente associadas ao aumento da expressão de proteínas com função de defesa envolvidas na remoção de espécies reativas de oxigênio.
Tabela 4. Efeito de interação entre as médias da variável, concentração de proteínas das folhas e raízes de milho tratado com bioinsumos (Az, Azospirillum; ME, microrganismos eficientes; BsBm, Bacillus subtilis mais Bacillus megaterium; FNR + Az, fosfato natural reativo mais Az; FNR + BsBm; FNR + ME) ou não (TC, tratamento controle) e cultivado na presença (Capacidade de vaso 50%) ou não (Capacidade de vaso 100%) de restrição hídrica. Médias seguidas pela mesma letra maiúscula na horizontal dentro de cada condição/capacidade de vaso, e minúscula na vertical dentro de cada condição. Bioinsumos não diferem entre si pelo teste Tukey a p ≤ 0,05.
| Bioinsumos | Folhas (mg mL-1) | Raízes (mg mL-1) | x̅ geral (100%:50%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 100% | 50% | 100% | 50% | |||||||
| Valor | Sig. | Valor | Sig. | Valor | Sig. | Valor | Sig. | Folhas | Raízes | |
| TC | 1,971 | a A | 1,830 | a A | 2,10 | a B | 28,08 | a A | 1,90 | 15,09 g |
| Az | 1,431 | ab A | 1,959 | a A | 5,40 | a B | 30,51 | a A | 1,69 | 17,94 f |
| ME | 0,930 | b B | 1,911 | a A | 11,34 | a B | 37,77 | a A | 1,42 | 24,57 e |
| BsBm | 1,401 | ab A | 1,740 | a A | 14,64 | a B | 42,39 | a A | 1,57 | 28,53 d |
| FNR + Az | 1,740 | ab A | 2,061 | a A | 19,71 | a B | 46,35 | a A | 1,90 | 33,03 c |
| FNR + BsBm | 1,911 | ab A | 0,360 | b B | 21,96 | a B | 48,57 | a A | 1,13 | 35,25 b |
| FNR + ME | 1,959 | a A | 1,830 | a A | 24,57 | a B | 51,63 | a A | 1,89 | 38,10 a |
| Média | 1,740 | 1,830 | 14,25 B | 40,77 A | ||||||
| CV (%) | 24,056 | 3,554 | ||||||||
Sob condições de déficit hídrico e limitação nutricional, como a baixa disponibilidade de fósforo, plantas podem apresentar alterações na expressão proteica, refletindo ajustes metabólicos associados à adaptação ao estresse (ZHANG et al., 2022). Nesse contexto, pode ocorrer tanto a supressão quanto a estimulação da biossíntese de proteínas envolvidas em processos de defesa e manutenção celular (MA; DIAS; FREITAS, 2020; MANNINO et al., 2020). Os microrganismos promotores do crescimento de plantas (MPCP) modulam a síntese proteica e ativam enzimas as quais destoxificam peróxidos em plantas estressadas, ajudando-as a responder adaptativamente a homeostase celular (JOSHI et al., 2016). Portanto, os dados do presente estudo mostraram que a aplicação dos MPCP associado ou não ao FNR impactaram na concentração de proteínas solúveis em milho sob diferentes condições hídricas, com destaque para o papel crucial dos bioinsumos na resposta adaptativa das plantas ao déficit hídrico. Os dados sugerem que os bioinsumos melhoram a capacidade das plantas em manter a homeostase celular sob estresse, evitando danos nas biomoléculas. O aumento de 186% na concentração de proteínas solúveis nas raízes sob restrição hídrica reforça a hipótese de que a síntese proteica é intensificada para estabilizar membranas celulares e facilitar a sinalização associada ao estresse (SHARMA et al., 2019). Portanto, a aplicação dos bioinsumos é uma estratégia eficaz para mitigar os efeitos negativos do déficit hídrico, promovendo uma resposta adaptativa robusta e eficaz no milho. Ademais, análises morfofisiológicas conduzidas no mesmo delineamento experimental demonstraram que a inoculação com bioinsumos favoreceu a manutenção de parâmetros fisiológicos, incluindo trocas gasosas, bem como incremento na altura das plantas sob condições de déficit hídrico (SANCHES et al., 2025), fornecendo suporte complementar aos efeitos metabólicos observados no presente estudo.
Antioxidantes são mobilizados nas células vegetais como mecanismo de defesa contra os efeitos deletérios das espécies reativas de oxigênio (ERO), cujas concentrações tendem a aumentar sob variados estresses ambientais (MITTLER et al., 2022). Os sistemas de defesa antioxidante em plantas são classificados como enzimáticos e não enzimáticos e têm como objetivo manter a homeostase durante condições de estresse (BHAT et al., 2022; SOARES et al., 2019). Esses sistemas são fundamentais para remover as espécies reativas de oxigênio (ERO), que podem causar danos oxidativos a proteínas, lipídios, organelas, DNA e RNA (MITTLER, 2017; XING et al., 2023). A atividade antioxidante aumenta em plantas sob condições de estresse, tal como o déficit hídrico, demonstrando que a resposta antioxidante favorece a capacidade das plantas para superar esses estresses e melhorar seu desempenho (BHAT et al., 2022; NADARAJAH, 2020). Entre as enzimas envolvidas na degradação de ERO e que compõe o sistema de defesa antioxidante estão a catalase (CAT), a ascorbato peroxidase (APX), a superóxido dismutase (SOD), a guaiacol peroxidase (GPOD), entre outras (SOARES et al., 2019). As peroxidases, pertencentes ao grupo das oxidorredutases, catalisam diversas reações oxidativas em plantas, utilizando peróxido como substrato ou, em certos casos, oxigênio como aceptor de hidrogênio (MITTLER et al., 2022).
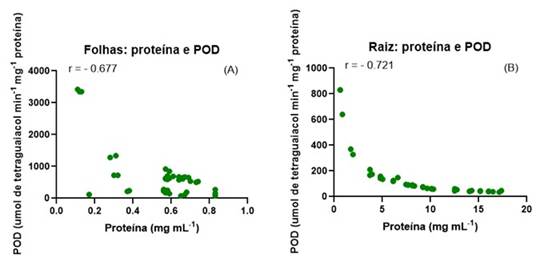
Os dados referentes ao desdobramento da atividade da gpod estão apresentados na tabela 5. Os tratamentos com bsbm ou fnr+az ou fnr+me induziram uma redução de aproximadamente 30% na atividade da enzima gpod em folhas de milho cultivado sem déficit hídrico (100% cp). Por outro lado, em plantas cultivadas com déficit hídrico (50% cp) os níveis da atividade da gpod foram incrementados com a aplicação de todos os bioinsumos, sendo o valor observado para o tratamento fnr+bsbm 29 vezes maior se comparado com o tratamento controle (50% cp). Isso indica que os bioinsumos aplicados melhoraram a eficiência dos mecanismos de defesa antioxidante das plantas, reduzindo a necessidade de alta atividade de gpod do milho cultivado sem déficit hídrico e aumentando a atividade dessa enzima sob déficit hídrico. Por outro lado, sob condições de déficit hídrico, os bioinsumos incrementam a atividade da gpod aumentando a capacidade de remoção dos oxidantes gerados durante o estresse. Tais dados não são corroborados nos níveis de proteínas (tabela 4), as quais seguiram um padrão diferente daquele observado para a atividade da gpod (tabela 5). Além disso, as análises de correlação de pearson mostraram correlações significativas negativas entre gpod e proteínas tanto para as folhas (-0,677) quanto para as raízes (-0,721) (figura 1). Isso sugere que as proteínas das raízes induzidas sob déficit hídrico, são proteínas de defesa antioxidante não determinadas neste estudo, como catalase e ascorbato peroxidase.
Tabela 5. Atividade da enzima Guaiacol Peroxidase – GPOD – das folhas e raízes de milho tratado com bioinsumos (Az, Azospirillum; ME, microrganismos eficientes; BsBm, Bacillus subtilis mais Bacillus megaterium; FNR + Az, fosfato natural reativo mais Az; FNR + BsBm; FNR + ME) ou não (TC, tratamento controle) e cultivado na presença (Capacidade de vaso 50%) ou não (CP 100%) de restrição hídrica. Médias seguidas pela mesma letra maiúscula na horizontal dentro de cada condição/capacidade de vaso, e minúscula na vertical dentro de cada condição. Bioinsumos não diferem entre si pelo teste Tukey a p ≤ 0,05.
| Bioinsumos | GPOD (µmol de tetraguaiacol min-1 mg-1 proteína) | |||||||
|---|---|---|---|---|---|---|---|---|
| Folhas | Raízes | |||||||
| 100% | 50% | 100% | 50% | |||||
| TC | 618,13 | ab A | 117,12 | c B | 766,72 | a A | 70,67 | a B |
| Az | 846,12 | a A | 606,43 | b A | 355,97 | b A | 60,37 | a B |
| ME | 928,19 | a A | 592,67 | b B | 183,42 | c A | 53,04 | a B |
| BsBm | 176,43 | c B | 784,99 | b A | 145,02 | cd A | 43,66 | a B |
| FNR + Az | 214,02 | c B | 610,27 | b A | 112,73 | cd A | 41,40 | a B |
| FNR + BsBm | 174,57 | c B | 3372,78 | a A | 109,31 | cd A | 39,93 | a B |
| FNR + ME | 240,18 | bc B | 687,96 | b A | 86,45 | d A | 38,34 | a A |
| CV (%) | 21,81 | 30,64 | ||||||
Assim, antioxidantes não enzimáticos e osmólitos são produzidos pelas plantas como parte do processo de defesa e remoção dos oxidantes danosos aos componentes celulares produzidos durante o déficit hídrico. Dentre os antioxidantes está a prolina, um aminoácido que além de antioxidante possui propriedades de osmoproteção (ZHANG et al., 2025). A prolina desempenha um papel vital nas plantas expostas a estresses extremos, como seca e salinidade e, atua como um dos principais constituintes das proteínas da parede celular, conferindo resistência devido à sua estrutura (ZHANG et al., 2025). A prolina desempenha um papel vital nas plantas expostas a estresses extremos, como seca e salinidade e, atua como um dos principais constituintes das proteínas da parede celular, conferindo resistência devido à sua estrutura. O grupo α-amino é uma amina secundária que proporciona rigidez conformacional excepcional às proteínas, fundamentais no processo de lignificação dos ramos (KAVI KISHOR, 2015), além de atuar no ajustamento osmótico e favorecer o aumento da tolerância na planta quando expostas a certos níveis de estresse, como o déficit hídrico (ZULFIQAR; ASHRAF, 2023).
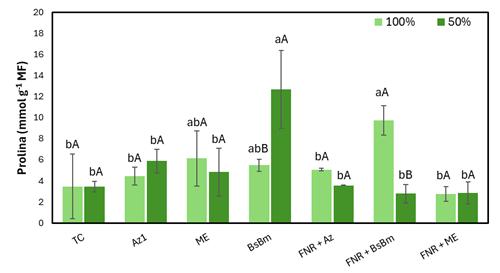
No presente estudo, a comparação entre as médias da concentração de proliomena em folhas de milho tratadas com bioinsumos e cultivadas sob déficit hídrico ou não, são ilustrada na Figura 02. Os resultados mostraram que, nos tratamentos sem déficit hídrico, as variações na concentração de prolina em relação ao tratamento controle (TC) foram significativas apenas no tratamento utilizando FNR+BsBm (≈ 9,0 µmol g⁻¹ MF) em comparação ao controle (TC ≈ 3,5 µmol g⁻¹ MF), sendo este o único a diferir significativamente, conforme indicado pelas letras associadas às médias (Figura 2). Sob condições de déficit hídrico, o tratamento BsBm apresentou maior concentração de prolina (≈ 12,5 µmol g⁻¹ MF) em relação ao controle (≈ 3,5 µmol g⁻¹ MF), também evidenciado pelas diferenças nas letras. Em relação às condições (100 e 50% CP) testadas, os tratamentos com BsBm (≈ 6,0 vs 12,5 µmol g⁻¹ MF) e FNR+BsBm (≈ 9,5 vs 2,5 µmol g⁻¹ MF) diferiram significativamente, conforme indicado pelas letras maiúsculas associadas às médias. Por outro lado, o tratamento com FNR+BsBm induziu um incremento na concentração de prolina livre em plantas sem déficit hídrico quando comparado com o mesmo tratamento de plantas com déficit hídrico. O acúmulo de prolina é relatado na literatura como uma resposta ao estresse causado pela desidratação, especialmente em condições de limitação hídrica (ZULFIQAR; ASHRAF, 2023). No presente estudo, o acúmulo de prolina sob déficit hídrico foi verificado apenas no tratamento BsBm, sugerindo que a inoculação com Bacillus subtilis + Bacillus megaterium desempenhou papel determinante na ativação de mecanismos osmoprotetores em milho crioulo. Em contraste, a associação BsBm + FNR promoveu incremento de prolina apenas sob condição de 100% de capacidade de vaso, sem efeito adicional em déficit hídrico (50% da capacidade de vaso), indicando que a resposta esteve majoritariamente vinculada ao consórcio bacteriano. A redução nos níveis de prolina em alguns tratamentos sob estresse hídrico reforça a complexidade das interações entre bioinsumos e mecanismos de defesa, evidenciando que a modulação osmótica não foi generalizada, mas dependente da inoculação específica.
A peroxidação lipídica em tecidos vegetais, como as folhas e raízes de milho pode ser avaliada através do método TBARS (Thiobarbituric Acid Reactive Substances). Este protocolo é utilizado para avaliar a extensão da degradação oxidativa dos lipídios, proporcionando uma estimativa indireta dos danos celulares causados pelo estresse oxidativo. Alterações no sistema redox das células vegetais podem desencadear a produção de compostos como o peróxido de hidrogênio (H2O2), levando à peroxidação lipídica (AKTER et al., 2021). A falta de água no solo intensifica o estresse oxidativo e a produção de ERO, resultando em peroxidação lipídica e danos subsequentes às membranas celulares (SACHDEV et al., 2021). Diversos subprodutos que intensificam os danos oxidativos são gerados pela peroxidação lipídica, tal como o malondialdeído (MDA), sendo o principal e mais investigado. O MDA é amplamente utilizado como marcador para identificar a peroxidação lipídica. As ERO, incluindo o H2O2, podem se formar nos cloroplastos, onde o O2 gerado reage com elétrons excedentes formando O 2●− (FOYER; HANKE, 2022).
Os níveis de TBARS nas folhas de milho variaram de 6,16 a 9,67 nmol MDA g⁻¹ MF na condição de 50% e de 6,70 a 9,67 nmol MDA g⁻¹ MF na condição de 100% da capacidade hídrica, sem diferença significativa entre os tratamentos dentro de cada condição, conforme indicado pelas letras associadas às médias (Tabela 6). As diferenças observadas ocorreram entre as condições de irrigação em alguns tratamentos, evidenciando que a redução para 50% da capacidade de vaso resultou em menores valores em comparação a 100%. Além disso, para os níveis de TBARS das raízes, observou-se efeito significativo da condição hídrica, com valores médios de 2,01 nmol MDA g⁻¹ MF sob 100% da capacidade de vaso e 1,46 nmol MDA g⁻¹ MF sob 50%, conforme indicado pelo teste de Tukey (p ≤ 0,05) (Tabela 6). Diferenças entre as condições hídricas foram evidenciadas pelas letras associadas às médias, indicando maiores valores sob 100% em comparação a 50%. Esses resultados observados sugerem que, sob déficit hídrico, as plantas apresentaram menores níveis de peroxidação lipídica quando comparadas à condição de 100% da capacidade de vaso, evidenciado pelos menores níveis de MDA. Esse resultado indica que não houve intensificação do dano oxidativo às biomoléculas nas condições avaliadas. Esse comportamento é acompanhado pela média dos níveis de proteínas, a qual apresentou valores menores nas raízes de plantas não submetidas ao déficit hídrico. A concentração de malondialdeído (MDA) é um indicador de estresse oxidativo em plantas, resultante da peroxidação lipídica, processo no qual os lipídios das membranas celulares são danificados pela ação de radicais livres. Nava et al. (2021) observaram uma correlação entre o teor de prolina, APX e TBARS na parte aérea, em que o malondialdeído estaria relacionado à manutenção da atividade enzimática de Baccharis trimera (Less.). No presente estudo, o tratamento FNR + ME apresentou valores de TBARS nas folhas de 6,70 nmol MDA g⁻¹ MF sob 100% e 6,12 nmol MDA g⁻¹ MF sob 50% da capacidade de vaso, enquanto o controle (TC) apresentou 7,80 e 6,16 nmol MDA g⁻¹ MF, respectivamente. Apesar das diferenças numéricas, não houve diferença estatística entre os tratamentos dentro de cada condição hídrica, conforme indicado pelas letras associadas às médias (Tabela 6).
Tabela 6. Níveis de peroxidação lipídica (TBARS) em folhas e raízes de milho tratado com bioinsumos (Az, Azospirillum; ME, microrganismos eficientes; BsBm, Bacillus subtilis mais Bacillus megaterium; FNR + Az, fosfato natural reativo mais Az; FNR + BsBm; FNR + ME) ou não (TC, tratamento controle) e cultivado na presença (Capacidade de vaso 50%) ou não (CP 100%) de restrição hídrica. Médias seguidas pela mesma letra maiúscula na horizontal dentro de cada condição/capacidade de vaso, e minúscula na vertical dentro de cada condição. Bioinsumos não diferem entre si pelo teste Tukey a 95% de confiança (p ≤ 0,05). ns Não significativo pelo mesmo teste.
| Bioinsumos | TBARS (nmol MDA g-1 MF) | |||
|---|---|---|---|---|
| Folhas | Raízes | |||
| 100% | 50% | 100% | 50% | |
| TC | 7,80 ab A | 6,16 ab B | 2,23 | 1,54 |
| Az | 9,67 a A | 5,93 ab B | 1,83 | 1,71 |
| ME | 7,23 b A | 7,88 a A | 1,95 | 1,66 |
| BsBm | 8,49 ab A | 6,93 ab B | 2,24 | 1,38 |
| FNR + Az | 7,50 b A | 7,91 a A | 1,85 | 1,09 |
| FNR + BsBm | 7,76 ab A | 5,22 b B | 1,89 | 1,22 |
| FNR + ME | 6,70 b A | 6,12 ab A | 2,09 | 1,66 |
| Média | 7,76 | 6,16 | 2,01 A | 1,46 B |
| CV (%) | 11,15 | 12,75 | ||
A associação entre fosfato natural reativo e microrganismos estimulou o aumento de proteínas radiculares e a estabilidade dos níveis de MDA, indicando ativação do sistema de defesa e preservação das membranas sob restrição hídrica. Esses efeitos concentraram-se no sistema radicular, evidenciando a modulação do metabolismo bioquímico por microrganismos promotores de crescimento em plantas de milho crioulo, independentemente do regime hídrico.
A UFFS pelo auxílio financeiro e bolsas concedidas. A FAPERGS pela bolsa concedida.
ABDELAAL, K.; ALKAHTANI, M.; ATTIA, K.; HAFEZ, Y.; KIRÁLY, L.; KÜNSTLER, A. The role of plant growth-promoting bacteria in alleviating the adverse effects of drought on plants. Biology, v. 10, n. 6, p. 520, 2021. 10.3390/biology10060520
AKTER, M. M.; IQBAL, M. T.; BASIRUN, W. J.; ALI, M. B.; AZAD, M. O. I.; SHAFIQ, S.; TAREQ, M. Z.; HOSSAIN, M. A. Role of reactive oxygen species and antioxidant defense in plants under abiotic stress. Frontiers in Plant Science, v. 12, 2021. 10.3389/fpls.2021.772018
BATES, L. S.; WALDREN, R. P.; TEARE, I. D. Rapid determination of free proline for water-stress studies. Plant and Soil, v. 39, n. 1, p. 205–207, ago. 1973. 10.1007/BF00018060
BHAT KA; MAHAJAN R; PAKHTOON MM; URWAT U; BASHIR Z; SHAH AA; AGRAWAL A; BHAT B; SOFI PA; MASI A; ZARGAR SM. Low temperature stress tolerance: an insight into the omics approaches for legume crops. Frontiers in Plant Science, v. 13, 3 jun. 2022. 10.3389/fpls.2022.888710
CONAB. Safra de grãos 2023/2024 está estimada em 294,1 milhões de toneladas. 2023. Disponível em: https://www.conab.gov.br/ultimas-noticias/5478-safra-de-graos-2023-2024-esta-estimada-em-294-1-milhoes-de-toneladas. Acesso em: 27 fev. 2026.
CONTINI, E.; MOTA, M. M.; MARRA, R.; BORGHI, E.; MIRANDA, R. A. de; SILVA, A. F. da; SILVA, D. D. da; MACHADO, J. R. de A.; COTA, L. V.; COSTA, R. V. da; MENDES, S. M. Série desafios do agronegócio brasileiro (NT2): milho – caracterização e desafios tecnológicos. EMBRAPA, 2019.
EI-MOSHATY, F. I. B.; PIKE, S. M.; NOVACHY, A. J.; SEHGAL, O. P. Lipid peroxidation and superoxide production in cowpea (Vigna unguiculata) leaves infected with tobacco ringspot virus or southern bean mosaic virus. Physiological and Molecular Plant Pathology, v. 43, n. 2, p. 109–119, 1993. 10.1006/pmpp.1993.1044
FOYER, C. H.; HANKE, G. ROS production and signalling in chloroplasts: cornerstones and evolving concepts. The Plant Journal, v. 111, n. 3, p. 642–661, 28 ago. 2022. 10.1111/tpj.15856
GENG, T.; JIA, F.; CAI, W.; WU, L.; GAN, B.; JING, Z.; LI, S.; MCPHADEN, M. J. Aumento da ocorrência de eventos consecutivos de La Niña sob o aquecimento global. Nature, v. 619, p. 774–781, 2023. 10.1038/s41586-023-06236-9
HELLUMS, D. T.; CHIEN, S. H.; PROCHNOW, L. I.; DEMISS, M.; SINGH, U. Cinquenta anos de pesquisa e desenvolvimento do IFDC sobre o uso de rocha fosfatada como fertilizante. Nutrient Cycling in Agroecosystems, v. 132, p. 15, 2026. 10.1007/s10705-025-10460-2
JOSHI, R.; WANI, S. H.; SINGH, B.; BOHRA, A.; DAR, Z. A.; LONE, A. A.; PAREEK, A.; SINGLA-PAREEK, S. L. Transcription factors and plants response to drought stress: current understanding and future directions. Frontiers in Plant Science, v. 7, 14 jul. 2016. 10.3389/fpls.2016.01029
KAVI KISHOR, P. B. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Frontiers in Plant Science, v. 6, 2015. 10.3389/fpls.2015.00544
KRUGER, N. J. The Bradford method for protein quantitation. In: Protein Protocols Handbook. New Jersey: Humana Press. p. 15–22. 10.1385/1-59259-169-8:15
KUMARI, S.; BASU, S.; KUMAR, G. A systematic review on the implications of concurrent heat and drought stress in modulating floral development in plants. Plant Science, v. 349, p. 112248, 2024. 10.1016/j.plantsci.2024.112248
LEITE, J. A.; MEIRA, A. L. Microrganismos eficientes: fundamentos e aplicações na agricultura sustentável. Recife: UFRPE, 2016.
LENGNICK, L. L.; KING, L. D. Comparação do status de fósforo dos solos gerenciados de forma orgânica e convencional. Revista Americana de Agricultura Alternativa, v. 1, n. 3, p. 108–114, 1986. 10.1017/S088918930000103X
MA, Y.; DIAS, M. C.; FREITAS, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Frontiers in Plant Science, v. 11, 2020. 10.3389/fpls.2020.591911
MANNINO, G.; NERVA, L.; GRITLI, T.; NOVERO, M.; FIORILLI, V.; BACEM, M.; BERTEA, C.M.; LUMINI, E.; CHITARRA, W.; BALESTRINI, R. Effects of different microbial inocula on tomato tolerance to water deficit. Agronomy, v. 10, n. 2, p. 170, 2020. 10.3390/agronomy10020170
MEENA, V. S.; MEENA, S. K.; VERMA, J. P.; KUMAR, A.; AERON, A.; MISHRA, P. K.; BISHT, J. K.; PATTANAYAK, A.; NAVA, S.; DOTANIYA, M. L.; DEEP, A.; SAXENA, A. K. Plant beneficial rhizospheric microorganism (PBRM) strategies to improve nutrients use efficiency: A review. Ecological Engineering, v. 107, p. 8-32, 2017. 10.1016/j.ecoleng.2017.06.058
MITTLER, R. ROS are good. Trends in Plant Science, v. 22, n. 1, p. 11–19, jan. 2017. 10.1016/j.tplants.2016.08.002
MITTLER, R.; ZANDALINAS, S. I.; FICHMAN, Y.; VAN BREUSEGEM, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology, v. 23, n. 10, p. 663–679, 2022. 10.1038/s41580-022-00499-2
NADARAJAH, K. K. ROS homeostasis in abiotic stress tolerance in plants. International Journal of Molecular Sciences, v. 21, n. 15, p. 5208, 2020. 10.3390/ijms21155208
NAVA, F. F. de M.; LUZ, V. C. da; STEMPKOWSKI, L. A.; GOTZ, L. F.; TONIAL, F.; CARGNELUTTI, D. Baccharis trimera (Less.) DC responses to water restriction. Revista Verde de Agroecologia e Desenvolvimento Sustentável, v. 16, n. 3, p. 229–237, 2021. 10.18378/rvads.v16i3.8360
NOVAIS, R. F.; ALVAREZ V., V. H.; BARROS, N. F.; FONTES, R. L. F.; CANTARUTTI, R. B.; NEVES, J. C. L. (Ed.). Fertilidade do solo. Viçosa, MG: Sociedade Brasileira de Ciência do Solo, 2007. 1017 p.
ROLLI, E.; MARASCO, R.; VIGANI, G.; ETTOUMI, B.; MAPELLI, F.; DEANGELIS, M. L.; GANDOLFI, C.; CASATI, E.; PREVITALI, F.; GERBINO, R.; CEI, F. P.; BORIN, S.; SORLINI, C.; ZOCCHI, G.; DAFFONCHIO, D. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environmental Microbiology, v. 17, n. 2, p. 316–331, 25 fev. 2015. 10.1111/1462-2920.12439
SACHDEV, S.; ANSARI, S. A.; ANSARI, M. I.; FUJITA, M.; HASANUZZAMAN, M. Abiotic stress and reactive oxygen species: generation, signaling, and defense mechanisms. Antioxidants, v. 10, n. 2, p. 277, 11 fev. 2021. 10.3390/antiox10020277
SANCHES, E. N. S.; SÉRGIO, B. M. S.; GENTILINI, G.; LIMA-MELO, Y.; CASTAMANN, A.; MAZIERO, S. M.; CARGNELUTTI, D. Landrace maize treated with bioinputs and natural phosphate tolerates water deficit. Research, Society and Development, v. 14, n. 7, e1514749170, 2025. 10.33448/rsd-v14i7.491702
SANTIAGO, G. M.; CARGNELUTTI, D.; CASTAMAM, A. Efficient microorganisms: an alternative tool for drought management in bean plants? Revista Verde de Agroecologia e Desenvolvimento Sustentável, v. 17, n. 1, p. 78-84, 2022. 10.18378/rvads.v17i1.9171
SARDANS, J.; MIRALLES, A.; TARIQ, A.; ZENG, F.; WANG, R.; PEÑUELAS, J. Growing aridity poses threats to global land surface. Communications Earth & Environment, v. 5, n. 1, p. 776, 2024. 10.1038/s43247-024-01935-1
SHARMA, A.; SHAHZAD, B.; REHMAN, A.; BHARDWAJ, R.; LANDI, M.; ZHENG, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules, v. 24, n. 13, p. 2452, 2019. 10.3390/molecules24132452
SOARES, C.; CARVALHO, M. E. A.; AZEVEDO, R. A.; FIDALGO, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environmental and Experimental Botany, v. 161, p. 4–25, 2019. 10.1016/j.envexpbot.2018.12.009
SOUSA, S. M.; GOMES, E. A.; SOUZA, F. A.; VASCONCELOS, M. J. V. Importância da morfologia radicular na eficiência da aquisição de fósforo. Embrapa, 2024.
VEJAN, P.; ABDULLAH, R.; KHADIRAN, T.; ISMAIL, S.; BOYCE, A. N. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules, v. 21, n. 5, p. 573, 2016. 10.3390/molecules21050573
VIEIRA, R. M. S. P.; TOMASELLA, J.; CUNHA, A. P. M. do A.; BARBOSA, A. A.; POMPEU, J.; FERREIRA, Y.; SANTOS, F. C.; ALVES, L. M.; OMETTO, J. P. Socio-environmental vulnerability to drought conditions and land degradation: an assessment in two northeastern Brazilian river basins. Sustainability, v. 15, n. 10, p. 8029, 2023. 10.3390/su15108029
XING, M.; LONG, Y.; WANG, Q.; TIAN, X.; FAN, S.; ZHANG, C.; HUANG, W. Physiological alterations and nondestructive test methods of crop seed vigor: a comprehensive review. Agriculture, v. 13, n. 3, p. 527, 2023. 10.3390/agriculture13030527
ZERAIK, A. E.; SOUZA, F. S.; FATIBELLO-FILHO, O.; LEITE, O. D. Desenvolvimento de um spot test para o monitoramento da atividade da peroxidase em um procedimento de purificação. Química Nova, v. 31, n. 4, p. 731–734, 2008. 10.1590/S0100-40422008000400003
ZHANG, H.; ZHU, J.; GONG, Z.; ZHU, J.-K. Abiotic stress responses in plants. Nature Reviews Genetics, v. 23, n. 2, p. 104–119, 2022. 10.1038/s41576-021-00413-0
ZULFIQAR, F.; ASHRAF, M. Proline alleviates abiotic stress induced oxidative stress in plants. Journal of Plant Growth Regulation, v. 42, n. 8, p. 4629–4651, 2023. 10.1007/s00344-022-10839-3
Esta obra está licenciada com uma Licença Creative Commons Atribuição 4.0 Internacional.